

{kind=link}
{kind=link}
{kind=link}
{kind=link}
冬凌草甲素协同TRAIL诱导OCM-1A细胞凋亡的作用机制
[查桂平 , 陆勤康, 陈剑英, 赖晓明]
, 陆勤康, 陈剑英, 赖晓明]
, 陆勤康, 陈剑英, 赖晓明]
|
|


第一作者:查桂平(ORCID:0000-0002-3301-931X),Email:13777068775@163.com
目的 通过实验研究考察肿瘤坏死因子相关的凋亡诱导配体(TRAIL)与冬凌草甲素联合使用时对脉络膜黑色素瘤细胞株OCM-1A细胞和C918细胞的抑制作用。方法 实验研究。以体外培养OCM-1A细胞和C918细胞为研究对象,加入不同浓度的冬凌草甲素(终浓度分别为0.625、1.25、2.5、5、10、20 μmol/L)和TRAIL(终浓度分别为6.25、12.5、25、50、100、200 ng/ml),利用MTT法检测细胞增殖活性,用PI单染结合流式细胞技术检测细胞周期分布,Annexin-V/PI双染检测细胞凋亡,用免疫印迹检测凋亡相关蛋白如死亡受体DR5、A-Caspase-3蛋白、XIAP蛋白表达量变化。数据采用单因素方差分析和独立样本 t检验进行分析。结果 TRAIL单独使用时,对OCM-1A细胞有一定程度的抑制,TRAIL浓度为200 ng/ml时抑制率最高为36.5%;冬凌草甲素单独作用时,对OCM-1A细胞的抑制不明显;当二者联合使用时,对OCM-1A的抑制作用明显增强,TRAIL 200 ng/ml联合冬凌草甲素20 μmol/L作用于OCM-1A时抑制作用最强,抑制率为87%,而对C918的抑制增强效果不明显。流式细胞仪检测结果发现处于G2期细胞显著增多,提示细胞被阻滞在G2/M期,免疫印迹结果显示DR5、A-Caspase-3表达增强,而凋亡抑制蛋白XIAP表达减弱。TRAIL与冬凌草甲素联合使用后对C918细胞的诱导凋亡作用不明显,差异没有统计学意义。结论 冬凌草甲素和TRAIL共同作用于OCM-1A细胞时,明显增强各自单独作用时候诱导的凋亡作用。
Objective: To study the inhibiting effect on OCM-1A cells and C918 cells from one of the choroidal melanoma cell lines by a combination of tumor necrosis factor related apoptosis inducing ligand (TRAIL) and Oridonin.Methods: In this basic experimental research, OCM-1A cells and C918 cells were cultured, and an experimental method was established to apply different concentrations of TRAIL (the final concentrations were 6.25, 1.25, 25, 50, 100, 200 ng/ml) and Oridonin (the final concentrations were 0.625, 1.25, 2.5, 5, 10, 20 μmol/L) to the OCM-1A and C918 cells. Then, the MTT method was used to detect cell proliferation activity. Cell cycle distribution and apoptosis were detected by flow cytometry, and the changes in apoptosis-related proteins such as DR5, a-Caspase-3 and XIAP were measured by Western blot. Data were analyzed using ANOVA and independent t test.Results: TRAIL had a certain degree of inhibition on OCM-1A cell proliferation. The highest inhibition rate was 36.5% when the concentration of TRAIL was 200 ng/ml. Inhibition of OCM-1A cell proliferation was not obvious with Oridonin. When TRAIL and Oridonin were simultaneously applied to OCM-1A cells, the inhibiting effect on OCM-1A was significantly enhanced. When TRAIL 200 ng/ml and Oridonin 20 μmol/L affected OCM-1A, the inhibition was the strongest, with an inhibition rate of 87%. The inhibitory effect on C918 was not significantly enhanced. The results of the cell cycle showed that TRAIL and Oridonin had an effect on OCM-1A cells with a significant blocking effect on G2/M. The results of Western blot showed that the expressions of DR5 and a-Caspase-3 were maintained, while the expression of apoptosis protein XIAP was suppressed. When TRAIL and Oridonin had a simultaneous effect on C918 cells, the apoptosis rate was not significantly different.Conclusions: The inhibiting effects of the combination of Oridonin with TRAIL on OCM-1A cells is significantly enhanced compared to the individual use of Oridonin or TRAIL alone.
脉络膜黑色素瘤(Choroidal melanoma, CM)是好发于成人的原发性眼部恶性肿瘤[1], 传统治疗方式以手术摘除患眼为主。近年来, 针对患者具体情况, 结合病变部位以及肿瘤大小采用不同的手段或者联合治疗已成为主要趋势。黑色素瘤的生物治疗是新的发展方向, 目前临床主要是应用大剂量干扰素, 多靶点激酶抑制剂以及抗血管内皮生长因子化合物进行治疗, 然而单药治疗效果往往不理想, 联合化疗可以显著提高疗效。肿瘤坏死因子相关的凋亡诱导配体(Tumor necrosis factor related apoptosis inducing ligand, TRAIL)是肿瘤坏死因子(Tumor necrosis factor, TNF)家族成员, 可通过作用于肿瘤细胞表面死亡受体(Death receptor, DR)来诱导多种肿瘤细胞系凋亡[2], 但也有部分细胞系由于DR缺失而对TRAIL耐受[3]。冬凌草甲素是双萜类生物碱, 具有抗氧化、抗炎作用, 同时具有抗肿瘤功效[4]。有研究发现冬凌草甲素对肝癌Hep G2细胞[5]、食管鳞状细胞癌EC9706细胞[6]、小鼠成纤维L929细胞[7]都有一定的诱导凋亡作用, 但具体作用机制尚不清楚。Paschall等[8]研究显示TRAIL与抗癌药物5-Fu联合使用时, 可以大量杀死结肠癌细胞系SW480细胞。同时有研究显示冬凌草甲素的类似物可以通过上调DR5诱导三阴乳癌细胞凋亡[9]。癌细胞具有高度的异质性, 也就意味着不同癌细胞亚群对同一种药物的敏感性不同。目前在临床上, 癌症治疗以多药联合应用为主。探索抗癌药物的联合应用将成为未来的抗癌治疗的趋势之一, 本研究主要探讨冬凌草甲素和TRAIL联合作用于CM时是否有一定的抑制效果, 并对其作用机制进行初步探索。
可溶性TRAIL蛋白购自上海近岸科技有限公司; 冬凌草甲素购自中国科学院成都生命研究所; MTT购自美国AMRESCO公司; RPMI1640购自美国Gibco公司; 胎牛血清购自美国GIBCO公司; DMSO、青霉素购自美国Life science公司; 紫杉醇购自上海源叶生物科技有限公司; 链霉素购自北京鼎国昌盛生物技术有限责任公司; DR5抗体、caspase-3抗体、X连锁凋亡抑制蛋白(X-linked inhibitor of apoptosis protein, XIAP)抗体、β -actin抗体购自德国Abcam公司。
常见的脉络膜黑色素瘤细胞OCM-1A和C918细胞株购自上海赛百慷生物技术公司, 细胞单层培养于含10%FBS、100 U/ml青霉素和100 U/ml链霉素的RPMI 1640的培养液中, 培养条件为37 ℃、5%CO2, 用0.25%胰酶消化并以1∶ 3~1∶ 5传代, 培养至对数生长期后用于实验。
冬凌草甲素组终浓度分别为0.625、1.25、2.5、5、10、20 μ mol/L; TRAIL组终浓度分别为6.25、12.5、25、50、100、200 ng/ml; 冬凌草甲素和TRAIL联合使用组终浓度分别为0.625 μ mol/L+6.25 ng/ml、1.25 μ mol/L+12.5 ng/ml、2.5 μ mol/L+25 ng/ml、5 μ mol/L+50 ng/ml、10 μ mol/L+100 ng/ml、20 μ mol/L+200 ng/ml。阴性对照组加入完全培养基来代替药物, 阳性对照组为5 μ mol/L紫杉醇。
收集处于对数生长期的OCM-1A和C918细胞, 调整细胞密度至5× 104/ml, 接种于96 孔培养板, 每孔100 μ l, 每个浓度设置3复孔, 细胞培养24 h后加入不同浓度的冬凌草甲素、TRAIL及冬凌草甲素+TRAIL处理细胞。药物作用48 h后每孔加20 μ l的5 mg/ml MTT, 37 ℃继续孵育4 h, 弃上清, 酶标仪检测各孔在560 nm 波长下的OD值, 按以下公式计算细胞抑制率, 细胞抑制率=(1-OD给药组/OD对照组)× 100%, 根据浓度-抑制率作图。以上实验重复3次。
1.5.1 流式细胞学检测细胞凋亡 取对数生长期细胞, 弃上清, 胰酶消化后以5.0× 104/ml接种于培养皿中, 依1.3中方法分组及加药, 设阴性对照组。培养48 h后取出细胞, 0.25%胰酶消化后以1 000 r/min离心10 min, 加PBS重悬后, 取一半转入EP管中, 剩余部分用于细胞形态分析。细胞悬液用PBS洗2次后加RNA酶置于37 ℃水浴30 min后, 加入1× 结合缓冲液300 μ l后, 重悬细胞, 分别加入PI染液和Annexin V-FITC染液各5 μ l, 室温避光孵育15 min, 加入200 μ l结合缓冲液, 转移至流式细胞上样管中, 于1 h内在流式细胞仪上检测, 分析各期细胞百分比。
1.5.2 细胞凋亡形态特征分析 方法1.5.1中取出的细胞悬液在1 000 r/min离心2 min后去上清液, 加入70%冷乙醇固定, 加入DAPI染液使其工作浓度为1 μ g/ml后染色2~3 min, 细胞悬液推片到载玻片上, 滤纸吸除多余水分, 立即置于荧光显微镜下观察细胞凋亡形态特征, 激发波长为360~400 nm。
收集阴性对照组及1.25 μ mol/L冬凌草甲素+12.5 ng/ml TRAIL, 2.5 μ mol/L冬凌草甲素+25 ng/ml TRAIL, 5 μ mol/L冬凌草甲素+50 ng/ml TRAIL处理48 h后的OCM-1A细胞, 加入RIPA细胞裂解液, 冰上裂5 min 后, 超声匀浆提取细胞总蛋白并定量。总蛋白经10% SDS-PAGE电泳分离后, 转移至 PVDF膜上, 室温下5%脱脂牛奶封闭1 h, 加入DR5、Caspase-3和XIAP一抗4 ℃过夜, 洗膜3次后, 加入辣根过氧化物酶标记的二抗, 室温孵育1 h, 再洗膜3次, 通过ECL试剂盒进行显影检测。β -actin为内参蛋白。
实验研究。采用SPSS 20.0统计软件进行分析。数据符合正态分布, 以均数± 标准差表示。多组间比较先分析方差齐性, 方差齐性者用ANOVA方差分析。2组间比较采用独立样本t检验。以P< 0.05为差异有统计学意义。
冬凌草甲素和TRAIL作用OCM-1A细胞48 h后对其增殖的抑制率见图1A, 冬凌草甲素和TRAIL作用C918细胞48 h后对其增殖的抑制率见图1B。TRAIL单独使用时, 对OCM-1A细胞有一定程度的抑制, 当TRAIL浓度为200 ng/ml时抑制率最高为36.5%; 当冬凌草甲素单独作用时, 对OCM-1A细胞的抑制不明显; 当二者联合使用时, 对OCM-1A的抑制作用显著增强, 当冬凌草甲素20 μ mol/L联合TRAIL 200 ng/ml作用于OCM-1A时抑制作用最强, 抑制率为87%, 显著高于20 μ mol/L冬凌草甲素(P=0.0036)和200 ng/ml TRAIL(P=0.0067)单独作用时的抑制作用。而冬凌草甲素联合TRAIL对C918的抑制效果的增强作用不明显。该结果提示OCM-1A细胞对冬凌草甲素和TRAIL的联合使用较C918细胞更为敏感, 后续实验选择OCM-1A作为主要的研究对象。
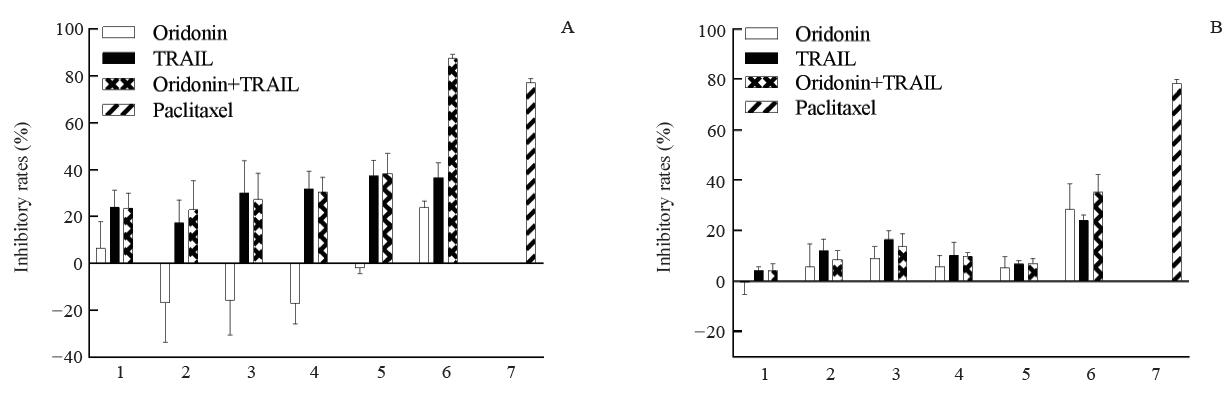 | 图1. 不同浓度冬凌草甲素和肿瘤坏死因子相关凋亡诱导配体(TRAIL)单独及联合作用对OCM-1A和C918细胞的抑制作用 A:OCM-1A细胞; B:C918细胞。1-6中冬凌草甲素浓度分别为0.625、1.25、2.5、5、10、20 μ mol/L, TRAIL浓度分别为6.25、12.5、25、50、100、200 ng/ml; 7为5 μ mol/L紫杉醇Figure 1. Inhibition of OCM-1A and C918 cells by different concentrations of Oridonin and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) alone or in combination. A: OCM-1A cells; B: C918 cells. The concentration of oridonin in 1-6 was 0.625, 1.25, 2.5, 5, 10, 20 μ mol/L, and the TRAIL concentrations were 6.25, 12.5, 25, 50, 100, 200 ng/ml, respectively; Group 7 indicated 5 μ mol/L of paclitaxel. |
冬凌草甲素和TRAIL联合作用于OCM-1A细胞后处于G2期的细胞有增多的趋势。对照组流式细胞显示为典型的G0/1、S、G2/M峰型。与阴性对照组比较, 冬凌草甲素组和TRAIL组的细胞周期变化无明显差异, 冬凌草甲素和TRAIL联合使用组G0/1期细胞的比例降低, G2/M期细胞随着药物浓度的增高也相应增多, 但处于S期的细胞比例与对照组相比无明显差异, 见图2。
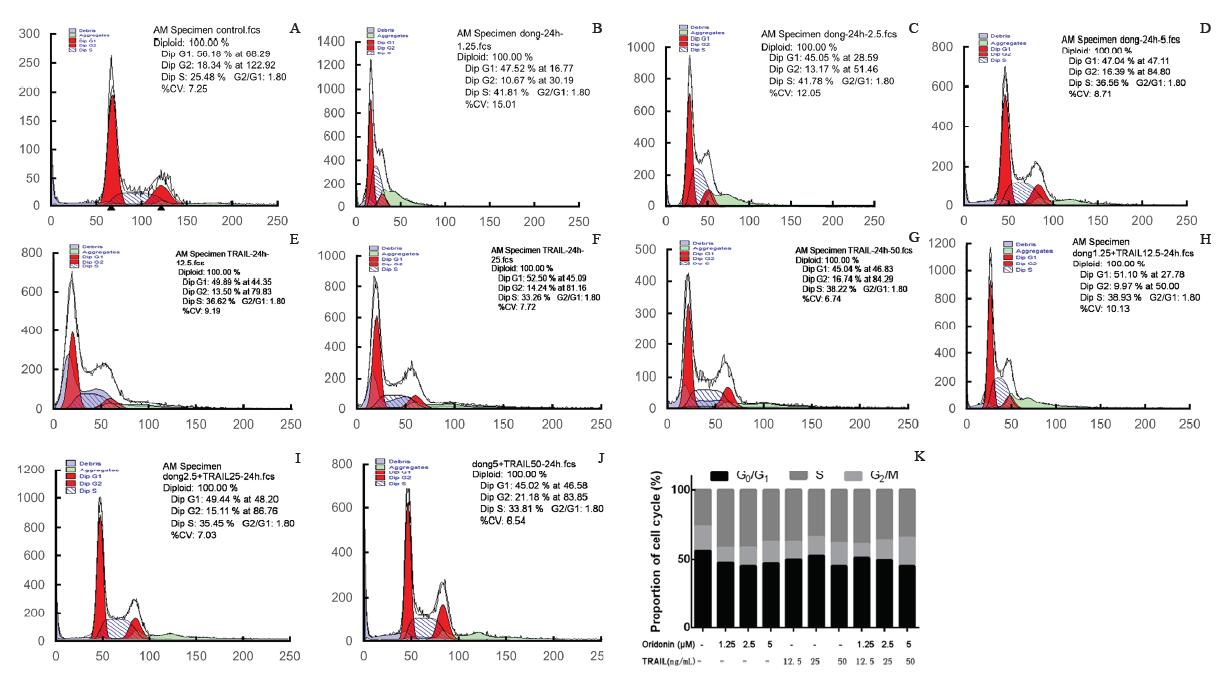 | 图2. 冬凌草甲素和肿瘤坏死因子相关凋亡诱导配体(TRAIL)单独或联合作用于OCM-1A细胞后细胞周期检测结果 A:正常对照组; B:1.25 μ mol/L冬凌草甲素组; C:2.5 μ mol/L冬凌草甲素组; D:5 μ mol/L冬凌草甲素组; E:12.5 ng/ml TRAIL组; F:25 ng/ml TRAIL组; G:50 ng/ml TRAIL组; H:1.25 μ mol/L冬凌草甲素和12.5 ng/ml TRAIL联合组; I:2.5 μ mol/L冬凌草甲素和25 ng/ml TRAIL联合组; J:5 μ mol/L冬凌草甲素和50 ng/ml TRAIL联合组; K:不同浓度冬凌草甲素和TRAIL作用于OCM-1A细胞后对细胞周期的影响Figure 2. The cell cycle detection of OCM-1A treated by Oridonin and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). A: Control; B: 1.25 μ mol/L Oridonin; C: 2.5 μ mol/L Oridonin; D: 5 μ mol/L Oridonin; E: 12.5 ng/mL TRAIL; F: 25 ng/mL TRAIL; G: 50 ng/mL TRAIL; H: 1.25 μ mol/L Oridonin+12.5 ng/mL TRAIL; I: 2.5 μ mol/L Oridonin+25 ng/mL TRAIL; J: 5 μ mol/L Oridonin+50 ng/mL TRAIL; K: The cell cycle effects of OCM-1A treated by Oridonin and TRAIL. |
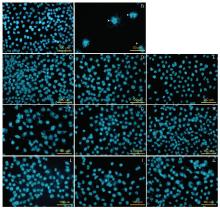
由图3可见, 阴性对照组细胞核比较完整, 无明显凋亡形态细胞可见。阳性对照组OCM-1A细胞出现典型的凋亡小体, 且无完整细胞核。1.25 μ mol/L冬凌草甲素组和2.5 μ mol/L冬凌草甲素组中没有出现大量凋亡细胞, 5 μ mol/L冬凌草甲素组出现了较多凋亡细胞。12.5、25、50 ng/ml TRAIL组中随着药物浓度升高, 细胞状态变差, 但尚未见大量凋亡细胞出现。1.25 μ mol/L冬凌草甲素+12.5 ng/ml TRAIL组出现较少的凋亡形态细胞, 2.5 μ mol/L冬凌草甲素+25 ng/ml TRAIL组和5 μ mol/L 冬凌草甲素+50 ng/ml TRAIL组可见大量凋亡小体出现, 提示细胞凋亡增多。可见随着上述2种药物浓度的增加, OCM-1A细胞的状态变差, 凋亡小体增加。且联合应用对OCM-1A细胞的凋亡作用要强于单独应用。
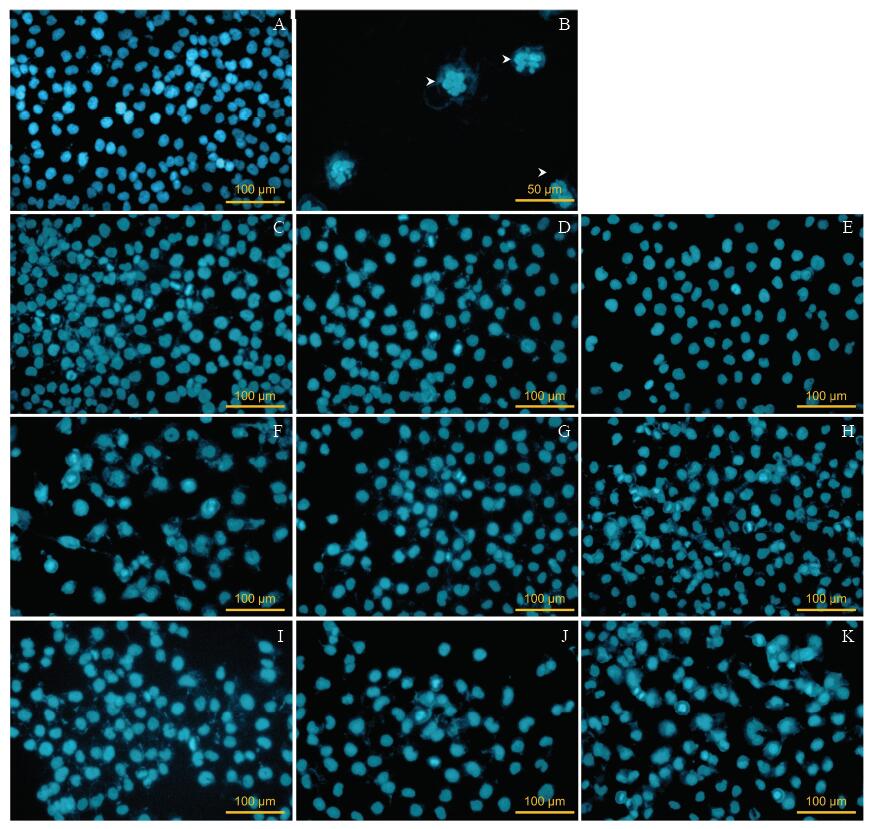 | 图3. 冬凌草甲素和肿瘤坏死因子相关凋亡诱导配体(TRAIL)单独或联合作用后OCM-1A细胞核的镜下形态特征 A:阳性对照组; B:5 μ mol/L紫杉醇组; C:1.25 μ mol/L冬凌草甲素组; D:2.5 μ mol/L冬凌草甲素组; E:5 μ mol/L冬凌草甲素组; F:12.5 ng/ml TRAIL组; G:25 ng/ml TRAIL组; H:50 ng/ml TRAIL组; I:1.25 μ mol/L冬凌草甲素+12.5 ng/ml TRAIL组; J:2.5 μ mol/L冬凌草甲素+25 ng/ml TRAIL组; K:5 μ mol/L冬凌草甲素+50 ng/ml TRAIL组Figure 3. Microscopic morphology of OCM-1A nuclei treated by Oridonin or tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). A: Control; B: 5 μ mol/L Paclitaxel; C: 1.25 μ mol/L Oridonin; D: 2.5 μ mol/L Oridonin; E: 5 μ mol/L Oridonin; F: 12.5 ng/mL TRAIL; G: 25 ng/mL TRAIL; H: 50 ng/mL TRAIL; I: 1.25 μ mol/L Oridonin+12.5 ng/mL TRAIL; J: 2.5 μ mol/L Oridonin+25 ng/mL TRAIL; K: 5 μ mol/L Oridonin+50 ng/mL TRAIL. |
检测OCM-1A中XIAP、DR5和A-Caspase-3蛋白本底表达以及2种药物联合应用后情况, 如图4所示, 在泳道1中OCM-1A中可表达XIAP, 而DR5、A-Caspase-3蛋白并不表达, 而当冬凌草甲素和TRAIL联合作用后, 这2种蛋白可随药物浓度的升高表达增强, 并随联合药物浓度增加而表达增强; 推测联合组能诱导DR5的表达, 并可活化Caspase-3(表现为A-Caspase-3表达增强), 继而诱导细胞凋亡发生。而XIAP蛋白表达却随2种药物浓度升高而下调。在小剂量(1.25 μ mol/L冬凌草甲素+12.5 ng/ml TRAIL)下就可以诱导DR5的表达, 继而实现相互的协同促进OCM-1A细胞凋亡。见图4。
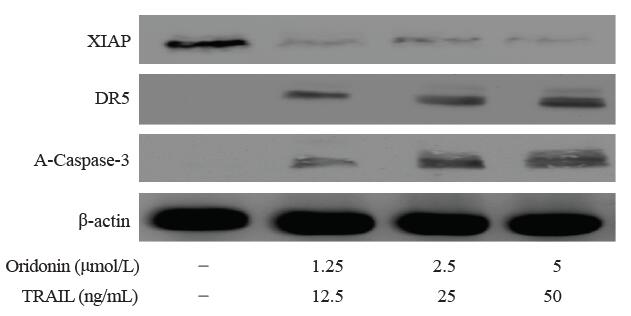 | 图4. 冬凌草甲素和肿瘤坏死因子相关凋亡诱导配体(TRAIL)联合作用OCM-1A细胞后凋亡相关蛋白表达情况Figure 4. Apoptosis-related protein expression of OCM-1A cells treated by Oridonin or tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). |
CM恶性程度高, 易侵袭转移, 预后极差, 严重威胁患者的健康, 治疗原则是“ 个性化” , 通常需要根据肿瘤的大小、位置、形态、生长速度等情况选用适合的治疗方法, 其中放疗效果较为良好, 但其局限性仍然存在。需综合考虑患者病情选择一套疗效更好, 复发率更低, 不良反应更少, 更能有效控制病情的化疗方案。
冬凌草甲素是中国自行研制的一种从唇形香茶属植物冬凌草中提取的一种以贝壳杉烯为骨架的四环二萜类化合物, 对多种肿瘤细胞都有一定的抑制作用[10]。Xiao等[11]在研究冬凌草甲素对非小细胞肺癌的作用时发现它可以通过提高凋亡相关蛋白BAX表达以及降低凋亡抑制蛋白BCL-2的表达, 从而促进肺癌细胞凋亡以及肺癌细胞的放疗敏感性。Zhu等[12]研究发现, 冬凌草甲素可抑制HepG2肝癌细胞增殖, 其机制可能为通过增加细胞周期相关蛋白, 如p-ATM、p-ATR、p-P53、p-Cdk1、p-CDC25C等, 从而发挥对肿瘤细胞的抑制作用。Gui等[13]研究冬凌草甲素对胰腺癌BxPC-3细胞的抑制作用时发现它能下调BxPC-3细胞中炎性因子相关的核转录因子的表达, 从而达到抗肿瘤作用。冬凌草甲素能够通过调控凋亡相关蛋白引起凋亡程序启动, 而使得细胞发生凋亡, 当冬凌草甲素引起周期相关蛋白表达异常时就可能通过阻滞细胞周期来使得细胞进入凋亡途径[14]。本研究发现, 冬凌草甲素和TRAIL联合作用OCM-1A细胞后, 处于G2期细胞显著增多, 提示细胞被阻滞在G2/M期, 免疫印迹结果显示DR5、A-Caspase-3表达增强, 而凋亡抑制蛋白XIAP表达减弱, 表明联合使用后可促进OCM-1A细胞凋亡。
自从TNF超家族被发现以来, 它的成员一直在增加, TRAIL就是其中一员, 研究人员通过搜索DNA数据库发现TRAIL与TNF和CD95L具有序列同源性并且它确实具有抑制肿瘤的功能, 从而将其归到TNF超家族当中来。因TRAIL能够选择性地靶向作用于肿瘤细胞表面的DR受体而对正常细胞不产生毒性, 对多种肿瘤都有一定的抑制作用[15], 然而在实际应用过程中发现许多类型肿瘤已经对其有耐受性[16], 这与肿瘤细胞表面的DR4、DR5功能缺失有关。肿瘤细胞内的自噬体对DRs的吞噬作用导致TRAIL耐药, 当使用3-Methyladenine(3-MA)破坏本底自噬体形成后, DR4/DR5释放到细胞膜表面, 就可增强TRAIL 耐药细胞对TRAIL的敏感性。
抗肿瘤物质往往只能对某种类型的肿瘤细胞起到较好的治疗效果, 并且这种效果可能由于使用时间的延长而发生肿瘤抑制效果下降的现象, 寻求新的用药手段就成为必然趋势。目前, 多种抗肿瘤物质联合使用已成为抗肿瘤治疗的一种新思路, 通过恢复抗肿瘤的信号通路而达到抑制肿瘤的目的, 其中紫杉醇联合顺铂新辅助化疗及放疗在早中期宫颈癌治疗中取得了比较好的效果[17]。TRAIL联合5-FU在体外抑制结肠癌SW480细胞在很早之前就有过报道[18]。最新研究报道将去氢骆驼蓬碱衍生物(B-9-3) (0.78~200.00 μ g/ml)与紫杉醇、春新碱、5-氟尿嘧啶、顺铂和表柔比星(0.78~200.00 μ g/ml)单独及联合作用于肺癌细胞LLC、结肠癌细胞HT-29、肝癌细胞HepG2和乳腺癌MCF-7 48 h后, 发现与B-9-3单独用药比较, 同浓度B-9-3与长春新碱和紫杉醇联合用药能够显著降低MCF-7和HT-29细胞的存活率, 因而认为B-9-3与不同抗肿瘤化合物联合具有协同作用, 提高细胞凋亡率[19]。该项研究推测肿瘤抑制物的作用机制是在同一信号通路的不同节点发挥作用而达到抑制肿瘤细胞增殖的目的。
本研究发现, TRAIL和冬凌草甲素一起作用时可以有效抑制原本对TRAIL耐受的OCM-1A细胞, 推测其作用机制可能是作用于OCM-1A细胞的自噬体, 使得肿瘤细胞表面的DR分子重新分布在细胞表面从而使得TRAIL发挥抗肿瘤效果, 以β -actin为内参, 测定凋亡抑制蛋白XIAP和凋亡蛋白家族Caspase成员A-Caspase-3的表达变化。
DR5是肿瘤细胞表面与TNF相关的凋亡诱导配体相结合的受体, TNF与之作用时可引发细胞进入凋亡途径, 在一些细胞的凋亡中发挥重要作用, 其高表达提示细胞对凋亡信号更加敏感, 而其低表达则为正常细胞或对肿瘤抑制剂耐受细胞。在本研究中, DR5在冬凌草甲素和TRAIL联合使用组中随药物浓度的升高表达增强, 推测上述药物可能通过DR5促进细胞的凋亡。
Caspase蛋白家族可通过特异性切割天冬氨酸的半胱氨酸蛋白酶而引起细胞凋亡, 凋亡为信号级联放大过程, 其中有多个Caspase家族蛋白参与其中, 如Caspase-8在细胞凋亡早期发挥作用, 对下游蛋白Caspase-3进行剪切得到剪切体A-Caspase-3, 在本研究中显示2种药物作用后可引起其高表达, 另外在本研究发现Caspase-3在联合给药组中出现活化, 并随联合组中2种药物浓度增加其活化程度而增强, 体现为剪切形式的Caspase-3表达水平随药物浓度呈现增加趋势; 另一类被称为执行者Caspases, 如A-Caspase-3主要通过剪切聚ADP核糖聚合酶(PARP), 进而引起DNA修复, 基因完整性监护功能降低, 受PARP负调控影响的Ca2+/Mg2+依赖性核酸内切酶的活性增高, 裂解核小体间的DNA, 引起细胞凋亡。正常情况下, Caspases以非活性酶原形式存在于细胞质中, 细胞接受凋亡信号后才会激活, 成为有促进细胞凋亡功能的蛋白质, 此时其表达含量尤其是终末效应分子A-Caspase-3表达含量明显上升。通过检测其表达含量可明确细胞是否发生大规模凋亡。XIAP是Peter等[20]于1996年在人的胚胎脑组织细胞中发现的IAP家族中的主要成员。XIAP基因位于X染色体q25, 其编码的蛋白含有497个氨基酸。人类XIAP是一种比较典型的IAPs, 由3个BIR结构域和3个RING结构组成, 这正是它能够抑制Caspase活性的结构基础。XIAP可以直接与激活的A-Caspase-3、Caspase-7和Caspase-9结合并抑制其活性, 进而抑制肿瘤细胞的凋亡[21], 通过检测其表达含量变化可为判定细胞凋亡是否增多提供支持。本研究中冬凌草甲素和TRAIL联合使用可活化Caspase-3, 免疫印迹检测Caspase-3蛋白治疗形式发现, 表达水平随2种药物联合作用的浓度增加而增加, 且XIAP蛋白表达随2种药物浓度升高而下调。提示可能通过抑制XIAP活化Caspase-3来诱导细胞凋亡。
利益冲突申明 本研究无任何利益冲突
作者贡献声明 查桂平:参与选题、实验设计及实验实施、论文撰写; 根据编辑部意见进行修改。陆勤康:参与选题、实验设计、修改论文的结果结论。陈剑英:参与选题、实验设计、实验结果分析。赖晓明:参与选题; 修改论文中关键性结果、结论; 根据编辑部修改意见进行核修
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|