{kind=link}
{kind=link}
{kind=link}
小鼠肌节同源盒基因同系物2条件性敲除在先天性白内障发生中的作用
[于紫燕, 于雯婷, 刘佳, 吴欣蔚, 赵江月, 张劲松 ]
]
]
|
|


目的 研究晶状体中肌节同源盒基因同系物2( Msx2)条件性基因敲除与先天性白内障发生的关系。方法 实验研究。选取条件性基因敲除小鼠Msx2CKO( Msx2fl/fl/Le- Cre+)为实验组,野生型小鼠Msx2WT( Msx2fl/fl)为对照组。取胚胎17.5 d(E17.5)Msx2WT小鼠胚胎头部组织作冰冻切片,采用RNA原位分子杂交方法检测 Msx2在眼组织内的正常表达。取2组小鼠E17.5和生后8 d(P8)眼球组织石蜡切片HE染色观察晶状体组织形态学变化。比较2月龄Msx2CKO和Msx2WT小鼠晶状体质量和直径,组间比较采用独立样本 t检验。结果 本研究观察到超过50%的2月龄Msx2CKO小鼠眼部出现角膜轻微混浊,晶状体变小(质量和直径),晶状体混浊及小眼球畸形。石蜡切片HE染色观察到Msx2CKO E17.5及P8小鼠晶状体内分化的纤维细胞排列紊乱,赤道部晶状体上皮细胞及邻近的纤维细胞中有空泡,排列明显紊乱。2月龄Msx2CKO组小鼠的直径小于Msx2WT组小鼠( t=4.80, P < 0.05),重量小于后者( t=14.29, P < 0.05)。结论 Msx2基因对小鼠晶状体发育起重要的调控作用,晶状体条件性敲除该基因可引起先天性白内障发生。
Objective: We investigated the relationship between msh homeobox 2 ( Msx2) conditional deletion and congenital cataract formation.Methods: In this experimental study, development of the ocular lens in Msx2CKO ( Msx2fl/fl/ Le- Cre+) and control Msx2WT ( Msx2fl/fl) embryos and postnatal mice were documented. Embryo heads at E17.5 (late embryo stage) were fixed, and frozen sections were processed for RNA in situ hybridization for detecting normal Msx2 expression. Morphological and histological differences were detected by hematoxylin and eosin staining of lenses from Msx2CKO and Msx2WT at E17.5 and P8 (8 days parturition). Lens axis diameter and weight were examined between Msx2CKO and Msx2WT mice at 2 months old. Statistical analyses were performed by independent t-tests.Results: More than 50% of the Msx2CKO mice exhibited minor corneal opacity, small lens formation (weight and axial diameter), nuclear sclerotic cataract, and microphthalmia compared with the wild-type littermates at two months. Msx2CKO embryos and postnatal specimens had differentiated lens fibers that were disorganized, and equatorial lens epithelial cells and lens fibers were vacuolated. Lens axis diameter in Msx2CKO mice was smaller than in Msx2WT mice, and weight was lighter in Msx2CKO mice than in Msx2WT mice.Conclusions: The Msx2 gene plays an important regulatory role during mouse lens development, and conditional deletion of Msx2 may cause congenital cataract formation.
先天性白内障是出生前即存在或出生后才逐渐形成的先天遗传或发育障碍的白内障, 其是一种较常见的儿童眼病, 是造成儿童失明和弱视的重要原因。先天性白内障的发病机制可分为遗传因素、环境因素以及原因不明三大类, 其中最重要的是遗传发育障碍等因素导致的先天性白内障。有关先天性白内障致病基因的研究很多, 已基本明确定位的先天性白内障相关基因主要包括四大类:①晶状体蛋白基因突变; ②膜蛋白基因突变; ③晶状体发育过程中的转录调节因子基因突变; ④细胞骨架蛋白基因突变[1]。其中, 晶状体发育过程中的转录调节因子基因突变是先天性白内障发生的主要原因之一, 已知的转录因子包括Pax6、Six3、Mab21L1等基因[2, 3, 4]。国内外研究证实肌节同源盒基因同系物2(Msx2)基因敲除小鼠会导致小眼球畸形, 晶状体、视网膜等眼组织发育严重异常, 但这些研究均采用的是传统Msx2基因全身敲除模型或转基因动物模型, 因此不能排除发育过程中眼组织间相互影响导致的继发晶状体发育异常[5]。Msx2基因对晶状体发育的调控作用还有待深入研究。本研究主要利用条件性敲除动物模型研究Msx2基因表达变化与晶状体发育及先天性白内障发生的关系。
选取中国医科大学实验动物中心提供的SPF级Msx2fl/fl小鼠[6]及Le-Cre转基因小鼠[7](Msx2+/+/Le-Cre+) (实验动物批准号:16005M)。将雌性Msx2fl/fl小鼠与雄性Msx2+/+/Le-Cre+小鼠交配, 获得实验组Msx2CKO(Msx2fl/fl/Le-Cre+)和对照组Msx2WT(Msx2fl/fl)胚胎及小鼠。动物的饲养与处理经中国医科大学动物伦理委员会审批通过(实验批准号:16005M), 并遵循眼科及视觉科学研究协会(The Association for Research in Vision and Ophthalmology)关于眼科和视觉研究中对动物的处理原则。
1.2.1 构建Msx2CKO动物模型及基因型鉴定 运用Cre-Loxp技术建立Msx2基因晶状体外胚层细胞特异性敲除小鼠模型。PCR确定胚胎及小鼠的基因型, 实验组Msx2CKO为Msx2fl/fl/Le-Cre+, 对照组Msx2WT为Msx2fl/fl。鼠尾PCR方法鉴定基因型[5]。Msx2 PCR 引物5'-GAA GTA GGC AGT CTC CAT CG-3' 和 5'-AAC ATC GCT CAG CCT GAA CC-3'。Cre knock-in的PCR引物为5'-CTC TGG TGT AGC TGA TGA TC-3'和5'-TAA TCG CCA TCT TCC AGC AG-3'。出生后2个月成鼠, 固定后裂隙灯显微镜下检查Msx2CKO和Msx2WT小鼠眼球发育情况。并解剖鼠眼, 取出晶状体, 观察Msx2CKO和Msx2WT小鼠晶状体的真实形态变化。
1.2.2 原位分子杂交 购买美国Roche公司T3、T7、SP6RNA聚合酶, 以体外PCR扩增的两端含有RNA聚合酶启动子的cDNA为模板, 利用RNA聚合酶体外转录合成地高辛标记的RNA探针。取Msx2WT胚胎17.5 d(Embryonic 17.5 d, E17.5)胚胎, 常规方法制作眼球组织冰冻切片。按照文献所提供的标准RNA原位分子杂交方法经预杂交、杂交、显色等步骤检测Msx2在眼组织内的正常表达[8]。
1.2.3 HE染色 分别选取Msx2CKO和Msx2WT同胞小鼠E17.5胚胎及生后8 d(8 days parturition, P8)小鼠, PCR确定基因型。胚鼠及小鼠采用二氧化碳窒息处死后(符合动物伦理), 取头部组织, 经固定、脱水、透明、包埋、切片等过程, 制作4 μ m厚平行视神经平面的整个眼球的石蜡切片, 进行常规HE染色。
1.2.4 晶状体测量 取Msx2CKO(n=28)和Msx2WT(n=14) 2月龄成鼠, 取出晶状体组织, 测量直径及称重。
实验研究。晶状体组织直径与重量数据结果以 ± s表示, 组间比较采用独立样本t检验。以P < 0.05为差异有统计学意义。
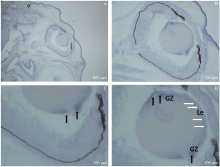
RNA原位分子杂交结果提示Msx2WT E17.5(n=4), Msx2基因在胚胎发育晚期的晶状体内有表达, 主要表达于晶状体上皮细胞尤其是晶状体赤道部的生发区(见图1)。
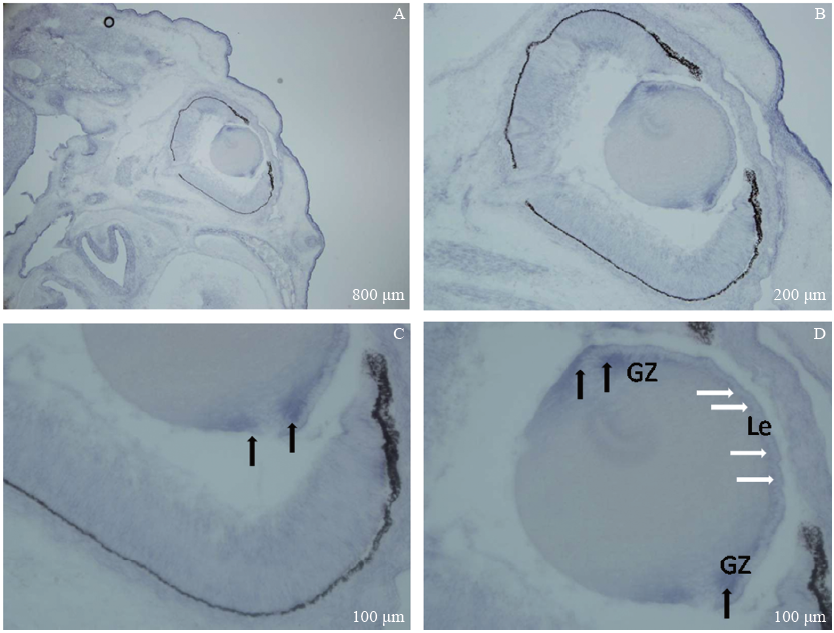 | 图1. 冰冻切片RNA原位分子杂交检测Msx2基因在Msx2WT胚胎发育晚期E17.5的晶状体内表达 A、B、C和D:Msx2主要表达于晶状体上皮细胞尤其是晶状体赤道部的生发区。Le:晶状体上皮细胞, 白色箭头示; GZ:生发区, 黑色箭头示Figure 1. In situ hybridization of Msx2 RNA in frozen sections of Msx2WT embryos at E17.5 (late embryo stage). A-D: Intense Msx2 hybridization signal is localized in the lens epithelium, especially the germinative zone near the equator. Le: Lens epithelial cells, white arrows; GZ: Germinative zone, black arrows. |
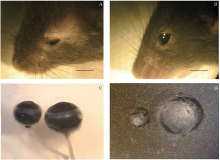
观察发现条件性敲除Msx2基因(n=12)后, 超过一半(7/12)的Msx2CKO小鼠前眼部表现为角膜轻微混浊, 晶状体变小, 可以观察到晶状体核性混浊(见图2), 大部分小鼠(10/12)发育为小眼球畸形。Msx2CKO(n=12)小鼠晶状体的体积明显小于Msx2WT小鼠(n=4), 并可观察到晶状体混浊的发生。
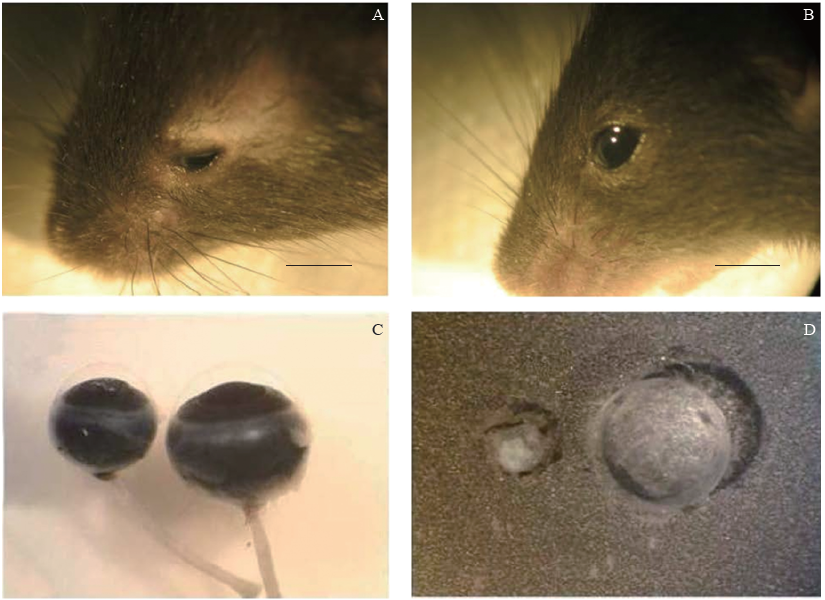 | 图2. 2月龄Msx2CKO与Msx2WT小鼠眼部表型 A:58.3% Msx2CKO 2月龄成鼠眼部表现为角膜轻微混浊, 晶状体变小, 可以观察到晶状体核性混浊, 小眼球畸形; B:Msx2WT小鼠2月龄成鼠表型, 没有发生白内障; C和D:解剖小鼠的眼球, 取出晶状体, 观察到Msx2CKO小鼠晶状体核性混浊, 且体积明显小于Msx2WT小鼠。标尺:1 cmFigure 2. Eye phenotype in Msx2CKO and Msx2WT mice at P2mo. A: More than 58.3% of the Msx2CKO mice exhibit minor corneal opacity, small lens formation, nuclear sclerotic cataract, and microphthalmia compared with the wild-type littermates (B) at 2 months. C, D: Enucleated eyes of Msx2CKO mice at 2 months of age are consistently microophthalmic, and the lenses are smaller than wild-type littermates. Scale bar: 1 cm. |
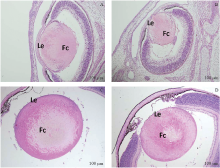
石蜡切片HE染色观察到在胚胎发育早期, Msx2CKO组小鼠眼部的发育异常并不十分明显, 各部分眼组织均有发育, 仅可观察到晶状体轻微发育变化。直至E17.5, 在表型明显的Msx2CKO胚胎中可以观察到晶状体未与角膜分离, 晶状体体积轻微变小, 晶状体内分化的纤维细胞排列紊乱, 赤道部晶状体上皮细胞及邻近的纤维细胞中有空泡, 排列明显紊乱。在出生后第8天, Msx2CKO组HE染色观察到晶状体上皮细胞的数量明显减少, 细胞排列紊乱, 在晶状体核部, 晶状体纤维排列紊乱, 出现少量空泡(见图3)。
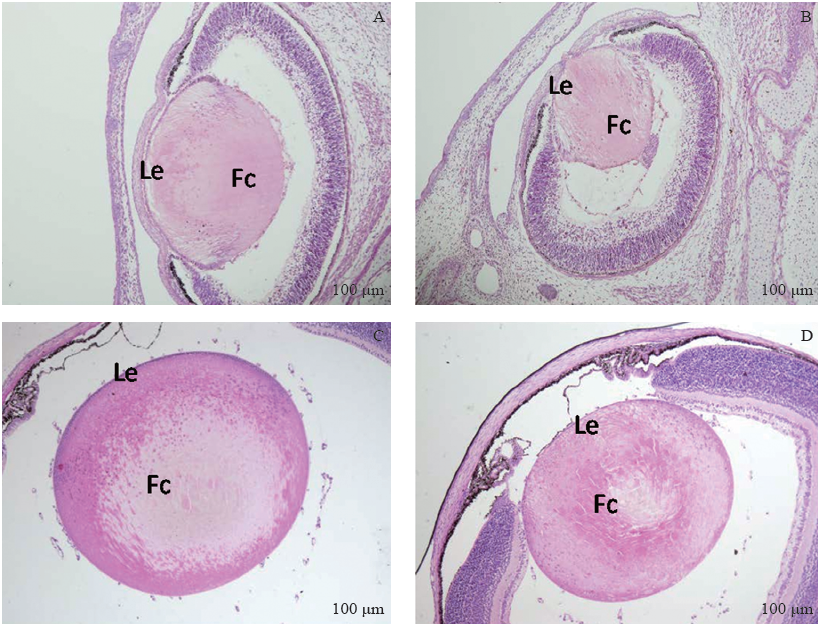 | 图3. E17.5和P8小鼠晶状体组织的形态学观察(HE染色) A:Msx2WT E17.5晶状体组织HE染色; B:在表型明显Msx2CKO胚胎E17.5石蜡切片HE染色中可以观察到晶状体未与角膜分离, 晶状体体积轻微变小, 晶状体内分化的纤维细胞排列紊乱, 赤道部晶状体上皮细胞及邻近的纤维细胞中有空泡, 排列显著紊乱; C:Msx2WT小鼠P8晶状体组织HE染色; D:Msx2CKO小鼠P8 HE染色可见晶状体上皮细胞的数量明显减少, 细胞排列紊乱, 在晶状体核部, 晶状体纤维排列紊乱, 出现少量空泡。Le:晶状体上皮细胞; Fc:晶状体纤维细胞Figure 3. Morphological and histological differences in lenses at E17.5 and P8. A: Hematoxylin and eosin (HE) staining of a wild-type control at E17.5. B: At E17.5 (late stage embryo). The lens is smaller and the persistent lens stalk is present in the severe type of Msx2CKO embryos. Differentiated lens fibers are disorganized, and the epithelial cells at the equator and the associated lens fibers are vacuolated. C: HE staining of a wild-type control at E17.5. D: At P8 (8 months parturition), lens epithelial cells are disorganized, with a clearly decreased in number. Differentiated lens fibers in the nucleus are disorganized and some are vacuolated. Le: Lens epithelial cell; Fc: Lens fiber cell. |
2月龄Msx2WT(n=14)小鼠与Msx2CKO(n=28)小鼠相比, 晶状体的质量减少37.2%, 直径减少30.7%。晶状体直径:Msx2WT组为(2.28± 0.39)mm, Msx2CKO组为(1.58± 0.47)mm, 组间差异有统计学意义(t=4.80, P < 0.05)。晶状体重量:Msx2WT组为(5.81± 0.27)mg, Msx2CKO组为(3.65± 0.53)mg, 组间差异有统计学意义(t=14.29, P < 0.05)。
先天性白内障是先天遗传或发育障碍的白内障, 可导致婴幼儿失明或弱视, 是一种严重的致盲疾病, 严重影响儿童的视力发育。随着眼科手术技术的发展, 先天性白内障的治疗效果虽然有显著的提高, 但是患儿还是要面对长期的弱视、眼球震颤、无调节力以及可能出现继发性青光眼等诸多问题。因此, 揭示先天性白内障的病因, 预防该疾病的发生有着重要的意义。
晶状体的发育及其中信号传导通路的形成是多阶段的过程, 包括了多种组织的相互作用。多种发育途径对晶状体的诱导发生和晚期的晶状体发育起重要的作用, 如BMP、FGF等信号传导通路等[9, 10]。同时一系列的转录因子也参与晶状体发育, 包括Pax6、Msx、Six3、Mab21L1、FoxE3、Prox1、Sox2、PitX3等。Msx2基因位于5号染色体长臂5q34-q35, 编码由263个氨基酸组成的蛋白质, 它可以与核心转录复合物结合调节转录。Msx2主要在脊椎动物的颅面部、四肢及乳腺等部位表达, 具有高度保守性, 它可以通过同源结构域与DNA序列(5'-AATTAG-3')相结合, 是新发现的眼发育调控基因[11, 12]。
国内外研究证实Msx2基因敲除小鼠会导致小眼球畸形, 晶状体、视网膜等眼组织发育严重异常。Msx2基因通过调控晶状体细胞的增殖、凋亡及晶状体上皮细胞内FoxE3基因的表达来调控晶状体早期发育[8]。这些研究均采用的是传统Msx2基因全身敲除模型或转基因动物模型, 虽然可以看到非常强烈的晶状体及视网膜发育障碍表现, 但是根据Ashery-Padan等[7]曾提出的晶状体-视网膜发育相互依存的重要理论, 证实晶状体与视网膜之间的发育存在相互诱导。因此, 已知的这些Msx2基因敲除后的表型变化可能是Msx2基因调控作用与组织发育诱导作用两者共同作用的结果。Msx2基因的真实功能还需要进一步的证明, Msx2基因对晶状体发育的调控作用还有待深入研究。
因此, 本研究构建了Msx2基因条件敲除小鼠模型, 仅在表面外胚层敲除了Msx2基因的表达, 对神经外胚层的Msx2基因无影响, 这样就避免了上面所说的那些问题。表面外胚层条件性敲除Msx2基因后, 眼部虽然也存在发育异常, 但是表型变化明显减轻, 小鼠的晶状体会明显变小, 并出现先天性白内障, 但是未观察到有小鼠出现无晶状体发育的情况。本研究观察了Msx2基因在晶状体发育晚期的正常表达, 发现Msx2在E17.5主要表达在晶状体上皮细胞中, 尤其是晶状体赤道部的细胞, 在分化的晶状体细胞中表达很少。而过去对晶状体发育早期的观察中发现Msx2基因最早在E9时就在表皮外胚层有表达, 随着发育表达在整个晶状体泡中, E12.5时在分化的晶状体细胞中表达强于前部晶状体上皮细胞。这说明Msx2基因随着晶状体的发育, 表达位置有所改变, 最终仅局限于有增殖能力的晶状体上皮细胞中。
本研究又观察了对应的同时期HE组织学染色结果发现晶状体纤维细胞排列紊乱, 赤道部晶状体上皮细胞及邻近的纤维细胞中有空泡。待出生后, 晶状体的组织学检查可以看到晶状体中央的纤维细胞排列紊乱, 有空泡等出现, 符合先天性白内障的组织学改变特点。
综上, 本研究主要通过构建表面外胚层条件性敲除Msx2基因的小鼠模型, 观察了该动物模型的表型特点, 发现晶状体发育受到影响, 同时出现先天性白内障的表型。先天性白内障的发病原因很多, 转录因子调控异常是导致先天性白内障的重要环节, Msx2基因也是先天性白内障发生的一个较重要的转录调控因子。
利益冲突申明 本研究无任何利益冲突
作者贡献声明 于紫燕:参与选题设计, 搜集数据, 资料的分析和解释; 撰写论文; 根据编辑部的修改意见进行修改。赵江月:参与选题设计, 撰写修改论文, 参与修改论文关键性结果、结论。刘佳, 吴欣蔚:参与搜集数据。张劲松:参与选题设计, 根据编辑部的修改意见进行核修
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|